Ліпіди
ЛІПІ́ДИ (від грец. λίπος — жир) — численна група різних за хімічним складом, структурою та функціями речовин, наявних у складі всіх живих організмів. Їх обʼєднують однакові фіз.-хім. властивості: вони нерозчинні у воді й ін. поляр. розчинниках, розчинні в неполяр. (гідрофоб.) рідинах (ефірі, хлороформі, бензолі, тетрахлоретані тощо), які використовують для екстракції Л. з біол. обʼєктів (тканин, крові та ін.). За хім. структурою більшість Л. є склад. естерами вищих карбон. (жирних) кислот та спиртів (гліцеролу, сфінгозину, холестеролу тощо). До складу багатьох склад. Л. входять також залишки фосфор. кислоти, азотистих сполук (холіну, етаноламіну), вуглеводів тощо. Фіз.-хім. та біол. властивості Л. визначає їхній жирнокислот. склад. Довжина вуглец. ланцюга, ступ. насиченості жирних кислот, що входять до складу природ. Л. (нейтрал. жирів (див. Жири), фосфоліпідів, сфінголіпідів тощо), зумовлюють їхню консистенцію (тверді, рідкі) і поверхн. активність, зокрема здатність до комплексоутворення з білками, утворення міцел (транспорт. ліпопротеїнів), бішарів матриксу біол. мембран. До Л. належать вищі жирні кислоти — монокарбон. кислоти аліфатич. ряду, природні з парним числом атомів вуглецю нерозгалуженої будови, насичені або ненасичені. В природі у вільному неетерифіков. стані жирні кислоти зустрічаються рідко, у людей переважно у плазмі крові. Вони входять до складу різних Л., утворюючи складні естери або аміди. Кількість атомів вуглецю в молекулі — не менше 6-ти, найпоширеніші в природі мають від 10-ти до 24-х. Насичені жирні кислоти в своєму складі не мають подвій. звʼязків. Заг. формула — СnH2nO2 або CH3(СН2)пCOOH, де n — кількість вуглец. атомів в аліфатич. ланцюгу. Чим більша кількість атомів вуглецю, тим вища температура плавлення. Найрозповсюдженіші насичені жирні кислоти: СН3(СН2)2СООН — масляна (бутанова), СН3(СН2)12СООН — міристинова (тетрадеканова), СН3(СН2)14СООН — пальмітинова (гексадеканова), СН3(СН2)16СООН — стеаринова (октадеканова). У складі ненасичених жирних кислот — один або декілька подвій. звʼязків у молекулі. Найважливішими є олеїнова (С18Н34О2) з одним подвій. звʼязком (С18 : 1), лінолева (С18Н30О2) з трьома подвій. звʼязками (С18 : 3), ліноленова (С18Н30О2) з трьома подвій. звʼязками в ін. положенні (С18 : 3), арахідонова (С20Н32О2) з чотирма подвій. звʼязками (С20 : 4). Олеїнова кислота міститься майже в усіх відомих жирах. Жирні кислоти нерозчинні у воді, але розчинні у водних розчинах лугів і аміаку. При цьому утворюються солі жирних кислот (мила). Калієві та натрієві солі жирних кислот розчинні у воді, солі важких металів — нерозчинні. Біполярність жирних кислот, особливо солей, зумовлює їхню поверхн. активність. Жирні кислоти хімічно інертні, крім ненасичених, що здатні відновлюватися за подвій. звʼязками. Насичені жирні кислоти тверді, ненасичені — рідкі. Найбільше значення для організму людини та вищих тварин мають лінолева, ліноленова й арахідонова кислоти. Перші дві в організмі людини не утворюються, арахідонова в невеликій кількості може утворюватися з лінолевої або ліноленової в процесі обміну речовин. Ці кислоти повинні обовʼязково потрапляти в організм з їжею у складі рослин. олій (їх часто обʼєднують під назвою вітаміни F). Вони беруть участь у біосинтезі ейкозаноїдів. Вищі жирні кислоти використовують у виробництві мила, синтетич. каучуку, гумових виробів, лаків, емалей, оліф тощо. Залежно від хім. структури компонентів Л. поділяють на прості — ацилгліцероли (нейтрал. жири), стероїди, цериди (воски), та складні — фосфоліпіди (гліцеролфосфоліпіди, сфінгофосфоліпіди), гліколіпіди (глікозилгліцероли, глікосфінголіпіди). Див. Табл. 1, 2, 3.

Ацилгліцероли — складні естери трьохатом. спирту гліцеролу і вищих жирних кислот (нейтрал. жири). Стериди — складні естери цикліч. спирту стеролу та вищих жирних кислот. Стероли є 3-гідроксипохідними стерану (циклопентанпергідрофенантрену).
Найрозповсюдженішим стеролом тварин. організму є холестерол, що входить до складу плазматич. мембран і є попередником у синтезі ін. стеролів та їхніх похідних (стероїдів).

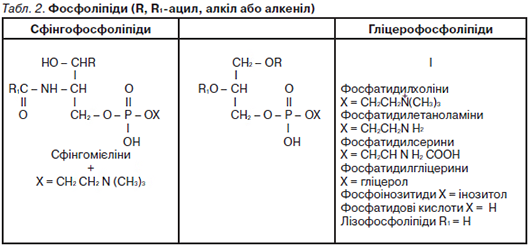
До біологічно важливих стеринів належать стероїдні гормони кори наднирк. залоз, чол. та жін. статеві гормони, вітаміни групи D та їхні похідні, жовчні кислоти. Рослини містять фітостерини. Цериди (воски) — прості Л., що є склад. естерами вищих жирних кислот і високомолекуляр. спиртів, зокрема цетилового (С16Н33ОН) та мірицилового (С30Н61ОН). До восків тварин. походження належать бджолиний віск, спермацет, ланолін, які використовують у фармації для виготовлення мазей, кремів, для виробництва косметич. засобів. До складу склад. Л. входять спирт (гліцерол, сфінгозин або інозит), фосфат, аміносполуки, вуглеводи. Складні Л. є поляр., амфіфільними сполуками. Більшість із них виконують структурні функції, входячи до складу біол. мембран. Численну групу склад. Л. становлять фосфоліпіди, які залежно від спирту у їхньому складі поділяють на гліцерофосфоліпіди (фосфогліцерини) та сфінгофосфоліпіди (фосфосфінголіпіди). Гліцерофосфоліпіди — складні естери гліцеролу та вищих жирних кислот, до їхнього складу входить також залишок фосфор. кислоти, етерифікований такими аміноспиртами, як холін, етаноламін і серин. Фосфодіефір. звʼязок у їхньому складі утворений гідроксил. групами холіну — фосфатидилхоліни (лецитини), етаноламіну — фосфатидилетаноламіни (кефаліни), серину — фосфатидилсерини, інозитоли (фосфатидилінозитоли). Сфінгофосфоліпіди — складні естери багатоатом. спирту сфінгозину та вищих жирних кислот, що містять також залишки фосфор. кислоти й аміноспиртів. У нерв. тканині людини та вищих тварин найпоширеніші сфінгомієліни (N-ацилсфінгенілфосфохоліни). Гліколіпіди — сполуки, у яких ліпідна частина ковалентно звʼязана з вуглеводною (глюкозою, галактозою та її похідними). Залежно від будови вуглевод. частини молекули їх поділяють на цереброзиди, гангліозиди, сульфотиди, що входять до складу мембран нерв. тканини. До Л. належать також біологічно активні похідні арахідонової кислоти — ейкозаноїди (простагландини, тромбоксани, лейкотриєни, ліпоксини), вищі спирти, ізопреноїди та їхні похідні; різноманітні стероїди, жиророзчинні вітаміни (А, D, Е, К). Ліпопротеїни — комплекси Л. різної хім. будови з білками, які є компонентами біол. мембран та знаходяться у плазмі крові людини. Ліпопротеїни плазми крові є молекуляр. комплексами різних Л. з білками, що утворюють міцелярні структури. За фіз.-хім. властивостями ліпопротеїни водорозчинні. Їхньою фізіол. функцією є міжорганне транспортування Л. Осн. ліпопротеїни плазми крові: хіломікрони — ультрамікроскопічні краплі жиру, вкриті шаром поверхнево актив. білків і фосфоліпідів; ліпопротеїни дуже низької щільності (β-ліпопротеїди); ліпопротеїни проміж. щільності; ліпопротеїни низької щільності (β-ліпопротеїни); ліпопротеїни високої щільності (α-ліпопротеїни). Ліпопротеїни крові — міцели, гідрофобне ядро яких складається з триацилгліцеролів і ефірів холестеролу, вкриті шаром білків і фосфоліпідів. Природні Л. виділяють з тварин. або мікроб. джерел, комбінуючи екстракцію орган. розчинниками з хроматогр. методами. Біол. функції Л. різноманітні. Нейтрал. Л. (жири) є формою депонування метаболіч. енергії. Фосфоліпіди, гліколіпіди і стерини — структурні компоненти біол. мембран; вони впливають на багато мембран. процесів, а саме на транспортування іонів і метаболітів, активність мембранозвʼязаних ензимів, міжклітинні взаємодії та рецепцію. Деякі гліколіпіди є рецепторами або корецепторами гормонів, токсинів, вірусів тощо. Фосфоінозитиди важливі у передаванні біол. сигналів. Ейкозаноїди — високоактивні внутр.-клітинні регулятори, міжклітинні медіатори та імуномодулятори, що беруть участь у розвитку захис. реакцій і запал. процесів. Деякі Л. є вітамінами та гормонами. Осн. масою Л., що потрапляють з тварин. і рослин. їжею в організм людини й тварин, є нейтрал. жири, а також незначна кількість фосфоліпідів та стероїдів. Розщеплення Л. відбувається в кишковику (дванадцятипалій кишці) за участі ензимів: ліпази, лецитинази, етаноламінфосфатази, гліцерофосфатази тощо. Суттєву роль у цьому процесі відіграють жовчні кислоти, які є емульгаторами Л. та активаторами ліпази. Нейтрал. жири в кишковику гідролізуються з утворенням ди-, моногліцеролів та вищих (карбон.) жирних кислот. Унаслідок гідролізу фосфатидилхоліну (лецитину) утворюються жирні кислоти, холін і фосфорна кислота. Під час гідролізу фосфатидилетаноламіну (кефаліну) вивільняється етаноламін (коламін). Ефіри холестеролу гідролізуються холестеролестеразою з утворенням вільних жирних кислот і холестеролу, циклічна будова якого в організмі не розщеплюється. Продукти гідролітич. розщеплення Л. та деяка кількість колоїд. часток емульгованого жиру всмоктуються ентероцитами кишковика в кровоносні судини (частково прямо, частково через лімфатичну систему) і транспортуються до різних органів та тканин, утворюючи жирові резерви, які використовуються відповідно до фізіол. потреб організму. Значна частина продуктів перетворення Л. у клітинах організму окислюється до кінц. продуктів обміну речовин. Деяка кількість гліцеролу і жирних кислот у стінках кишковика та ін. клітинах тканин ресинтезується з утворенням специфіч. для кожного організму жирів. Унаслідок врівноваженого надходження і витоку Л. у крові має місце їхня постійна концентрація — у середньому 500 мг (0,5 %), найбільше серед них фосфоліпідів (0,2 %), нейтрал. жирів (0,15 %), ефірів і вільного холестеролу (0,15 %). Гліцерол, який є продуктом гідролізу Л., внутрішньоклітинно ензиматично перетворюється на монофосфор. естер, потім на фосфогліцерин. альдегід (гліцерофосфат), наступне окислення якого відбувається за механізмом гліколізу. Вищі жирні кислоти (парні) окислюються переважно за механізмом β-окислення, внаслідок якого на кожному етапі відщеплюються 2 вуглец. атоми. Цей процес відбувається в матриксі мітохондрій, де окислюється вуглец. радикал жирної кислоти в другому (β) положенні відносно карбоксил. групи. Перед окисленням жирна кислота активується за участі коензиму А та АТР з утворенням ацил-КоА (ацил-похідного жирної кислоти): — R — СО — SКоА. Механізм β-окислення жирних кислот полягає в проходженні послідов. реакцій окислення і гідратації (Див. Табл. 4).
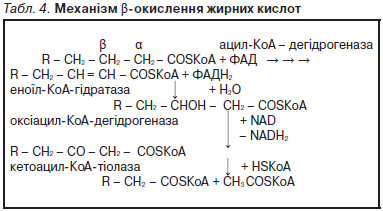
У результаті одного циклу β-окислення з молекули жирної кислоти вивільняється одна молекула ацетил-КоА, а вихідна молекула ацил-КоА скорочується на два вуглец. атоми і продовжує перетворення, поки не вивільниться остання молекула ацетил-КоА. Напр., β-окислення поширеної в природі пальмітинової кислоти має такий вигляд: С15 Н31 — СО — SКоА 7 циклів β-окислення 8 СН3 — СОSКоА. Ацетил-КоА окислюється в циклі трикарбон. кислот (циклі Кребса, лимонної кислоти). Вивільнена під час окислення жирних кислот енергія акумулюється в макроергіч. звʼязках АТР, на кожному етапі β-окислення утворюється 5 молекул АТР. У печінці існує фізіологічно важливий шлях утилізації ацетил-КоА, що призводить до утворення молекул альтернатив. метаболіч. палива, т. зв. кетонових (ацетонових) тіл, до яких належать ацетоацетат, гідроксибутират і ацетон, які використовують ін. тканини. Біосинтез жирних кислот відбувається переважно в цитозолі клітин, вихід. речовиною для нього є ацетил-КоА. Останній за участі біотинвмісного ензиму, СО2, АТР і Мn2+ утворює малоніл-КоА, який, конденсуючись з ацетил-КоА, перетворюється в ацетоацетил-КоА з вивільненням СО2. Завдяки реакціям дегідрування і декарбоксилювання утворюється бутирил-КоА, тобто відбувається додавання двовуглец. фрагмента до ацетил-КоА, і він перетворюється на чотиривуглец. радикал (С4). Згодом починається новий цикл реакцій, що полягають у подовженні вуглеводневого радикала карбон. кислоти ще на один двовуглец. фрагмент. Продуктом семи вказаних циклів є утворення радикала пальмітинової кислоти (С16). Синтез жирних кислот відбувається за участі мультиензим. комплексу синтетази жирних кислот, до складу якого входить декілька ензиматич. протеїнів. Центр. місце у цьому комплексі посідає ацилтранспортувал. протеїн (АТП), з яким сполучені всі ензиматичні білки, і до якого приєднуються ацильні похідні коензиму А. АТП має два SН-вмісні центри звʼязування: один — для акцептування ацетил-SКоА, другий (фосфорильований похідний вітаміну пантотенової кислоти) — для малонілового радикала. АТП виконує функцію кронштейна, який переносить проміжні продукти реакції від одного ензиму до іншого. Біосинтез нейтрал. жирів (триацилгліцеролів) відбувається ензиматично з відповід. ацилпохідних жирних кислот (ацил-КоА) та гліцерофосфату, в утворенні яких бере участь АТР. Ацетил-КоА (активна форма ацетату) може використовуватися у біосинтезі різних біологічно важливих сполук (стеролів, ацетилхоліну, ацетилглюкозаміну тощо). Здатність до синтезу холестеролу мають всі клітини тварин. походження, проте найбільша кількість ендоген. сполуки синтезується в печінці (50–80 %) та кишковику (10–15 %). Попередником у біосинтезі холестеролу є ацетил-КоА, який утворюється при окисленні глюкози або β-окисленні жирних кислот. Біосинтез холестеролу відбувається в цитозолі, складається з процесів епоксидації, окис. гідроксилювання, деметилювання і каталізується ензимами, що мають у структурі цитохром Р-450 та потребують наявності НАДРН2. У гепатоцитах із холестеролу синтезуються жовчні кислоти, які є гідроксильованими похідними холанової кислоти (холева, дезоксихолева, літохолева). З нього також синтезуються стероїдні гормони наднирк. залоз, статеві гормони, вітамін D3 та його похідні. Л. досліджують співроб. відділу біохімії ліпідів (Н. Гула) та лаб. технології біопрепаратів (З. Даценко) Інституту біохімії НАНУ (Київ). Зокрема у відділі розроблено і запатентовано біологічно активні сполуки з цитопротектор. дією — N-ацилетаноламіни (NАЕ); у лаб. одержано і запатентовано препарати з тканин гідробіонтів (мідії, рапани), до складу яких входять ω-3-ненасичені жирні кислоти, комплекс фосфоліпідів, плазмалогени тощо, для лікування гіпертонії, гормонал. порушень репродукт. системи, леген. захворювань.